摘要
传统的脑机接口解码皮层运动命令来控制外部设备。这些命令是高级认知过程的产物,发生在大脑区域的网络上,整合感觉信息,计划即将到来的运动动作,并监控正在进行的运动。我们回顾了最近在神经假体临床试验中在人类后顶叶皮层发现的认知信号。这些信号与在运动控制和身体监测的认知方面具有不同作用的皮质小区域一致,包括感觉运动整合、规划、轨迹表示、躯体感觉、动作语义、学习和决策。这些变量在相同的细胞群体中使用结构化的表示进行编码,这些表示结合了相关的感觉和运动变量,这种结构被称为部分混合选择性。不同的认知信号为传统的电机命令提供了补充信息,从而能够更加自然和直观地控制外部设备。
介绍
脑机接口(BMIs)已经显示出与瘫痪病人的人类大脑皮层接口来解码目的做简单的动作来控制外部设备,包括电脑光标和机器人。一般来说,录音是由初级运动皮层(M1)解码后的神经信号控制大脑的速度效应器。然而,其他大脑区域,在某种程度上甚至是运动皮层,编码与身体运动相关的高级认知信号(安德森等人,2010年).这些信号可以用来补充来自M1的简单移动命令,从而实现更复杂和高级的BMI(图1).身体质量指数范式还允许在皮层中发现具有科学意义的认知表征和功能。随着该领域的发展,更机械的认知知识将为更复杂的假肢提供科学框架。
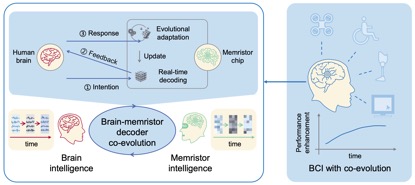
图1 认知身体质量指数示意图。翻译身体质量指数用户的解码意图,使他们能够与现实世界的物体互动,并通过新的感官信息更新他们的环境状态,这需要一个具有许多复杂部件的复杂设备。解码用户的高级认知目标(①),通过处理(④)PPC(②)中记录的神经活动电信号(③),身体质量指数向辅助设备(⑥)发送控制命令(⑤)。该设备装有传感器,将物理测量值(如压力、温度、本体感觉)传回身体质量指数(⑦)。这些信号被翻译成一种刺激模式(⑧),通过激活PPC中的神经元(⑨)来诱发自然的躯体感觉。图改编自安徒生(2019)。缩写:身体质量指数,脑机接口;后顶叶皮层。
在这篇综述中,我们使用术语BMIs这些设备也被称为脑机接口。在过去的10年中,神经技术领域的研究已经有了很大的发展,很难充分涵盖该领域的所有方面。如上所述,我们利用大脑皮层单个神经元的群体记录,致力于解决瘫痪问题。 从21世纪初开始,在非人类灵长类动物(NHPs)和人类中进行的运动控制BMI的首次尝试主要在M1(Ajiboye等人,2017年; Bouton等人,2016年; Carmena等人,2003年; 科林格等人,2013年; 弗里登伯格等人,2017年; Gilja等人,2015年; Hochberg等人,2006年, 2012; Pandarinath等人,2017年; Santhanam等人,2006年; Serruya等人,2002年; 泰勒等人,2002年; 韦斯伯格等人,2000年; Wodlinger等人,2014年).这个选择很有意义,因为M1是皮质下运动区和脊髓的皮质运动控制信号的主要来源。我们广泛研究了后顶叶皮质在我们实验室(Aflalo等人,2015年, 穆萨拉姆等人,2004年).我们推断,这个更高级的皮层区域涉及运动控制的更多认知方面,例如感觉运动转换,将具有重叠和互补的功能,有利于管理信息系统的运作。正如在这篇综述中看到的群体神经活动在人类的小块中,PPC编码许多变量,为身体质量指数应用提供丰富的信息库(Aflalo等人,2015年, 张等2017).这种高维度是由编码多个变量的单个神经元实现的,允许从PPC的小区域中读出过多的思想。 关于大脑皮层功能组织的研究历史已经在高度本地化的观点和更加分布式的观点之间转变。局部视图意味着皮质的解剖学上受限的区域负责特定的皮质过程(例如,用手抓握),而分布式视图意味着皮质的大部分或全部参与过程。在一个非常小的PPC区域内发现了许多认知表征,这使我们对关联皮层功能组织的理解转向了一个更为分散的观点。然而,正如我们将在这篇综述中看到的,PPC中的表示不是随机的,而是具有统计结构的。我们将这种结构称为部分混合选择性 (张等2017).此外,我们将看到初级区域仍然是更初级的,对它们的反应特性具有更有限的认知影响(贾法里等人,2020年).这些发现符合一个新兴的观点,即电路负责行为,皮层区域是一个或多个电路的节点。因此,皮质某个区域的记录反映了该区域的运作,以及它对回路其他节点的网络贡献。
脑机接口:从非人灵长类到人类
最初的身体质量指数实验是在NHPs(穆萨拉姆等人,2004年, Santhanam等人,2006年, Serruya等人,2002年, 泰勒等人,2002年).微电极阵列植入M1、PPC和背侧运动前皮质。这些动物被训练完成简单的运动任务,在这些任务中,它们用手的运动或伸展来控制计算机屏幕上光标的移动。计算机算法被训练使用神经群体活动来解码运动。最后,光标的控制从实际行为切换到解码的神经信号。NHPs将了解到他们不需要移动他们的四肢或手来完成任务,他们可以在精神上完成。在M1,主要的控制信号是效应器的预期速度。在PPC中,由解码器读出的信号更抽象,并指示运动的空间目标(穆萨拉姆等人,2004年).但是,PPC中还有一个轨迹信号可以用于轨迹解码。该PPC轨迹信号的动态稍微滞后于运动皮层轨迹信号的动态,并且这些动态与效应器的当前运动的状态估计相一致(Mulliken等人,2008年).状态估计是内部模型其捕获根据运动指令和感觉反馈信号计算的身体状态的当前估计。这种内部估计被认为占据了高阶运动区域,例如PPC。因此,这些NHP实验,尤其是PPC中的实验,与a认知神经修复术一种使用更抽象的预期目标和身体状态的内在表现。
下一个里程碑式的步骤是从BMI的动物模型转向人类临床试验(Aflalo等人,2015年; 巴特尔等人,2008年; 科林格等人,2013年; Hochberg等人,2006年, 2012) (图1).参与者瘫痪了,最常见的原因是高水平的脊髓损伤四肢瘫痪(四肢全部瘫痪),或部分瘫痪,来自肌萎缩侧索硬化的晚期。从转化的角度来看,人体临床试验是重要的一步。对于四肢瘫痪的受试者,许多人在进入临床研究之前已经有脊髓损伤多年。特别是对于运动皮层,不清楚长时间不用是否会影响神经元的反应特性。令人欣慰的是,运动皮层被发现对假肢控制足够完整(Hochberg等人,2006年).
使用神经信号控制设备的能力对瘫痪的人来说是一大进步。然而,瘫痪病人除了运动麻痹之外,通常还有明显的感觉缺陷。躯体感觉的丧失会带来直接和重大的健康挑战、使人虚弱的神经病和疼痛,以及基本身体功能如膀胱和肠道控制的困难。除了功能结果和生活质量的改善,感觉的恢复还可以改善对辅助设备的控制,如灵巧的机器人手(本斯梅亚&米勒2014, 韦伯等人,2012年).解决这个问题的一种方法是通过对体感皮层的微刺激来引入人工感觉(图1).由于来自人类参与者的诱发感觉的描述质量的口头报告,认知感觉BMI的发展最近已经加速。
对于认知运动BMI来说,转向人类已经打开了一个可能性的新领域(Aflalo等人,2015年).以前,很难知道NHPs如何执行身体质量指数控制任务。相反,人类有语言,可以报告他们控制身体质量指数的心理策略。此外,人类参与者可以被指导并且需要很少的训练来执行各种任务。因此,一系列认知变量可以在人类身上用最少的训练来测试。使用NHPs,探测认知需要为每项任务进行数月的训练。如此广泛的训练限制了大脑皮层某个区域的视野。一个主要的例子是外侧顶内区。这个区域是眼球运动皮层回路的一部分,它接收感觉输入,如来自纹外皮层的密集视觉投射。它投射到眼球运动区域,包括前视野和上丘。嘴唇在视觉运动整合和眼球运动中的明确作用使其成为研究通知眼球运动行为的高级认知过程的流行目标。经过几个月的训练后,根据任务的性质,单一的职能角色被分配到这一领域——注意力、计划、分类、物体识别和决策(比斯莱和戈德堡2003年, 弗里德曼和阿萨德2006, Gnadt & Andersen 1988年, 普拉特和格里姆彻1999年, 夏德伦和纽森1996年, Snyder等人,1997年).正如在“部分混合选择性”一节中所讨论的,LIP可能在所有这些功能中起作用。
如上所述,人类研究的一个优势是几乎不需要培训,将对现有功能组织的可能学习影响降至最低。与人类研究类似,NHPs的实验旨在了解皮质区域在感知、认知或运动方面的作用。假设是实验揭示了该区域的潜在功能。然而,NHPs中记录之前的广泛训练提出了一个问题,即学习在塑造被研究的神经回路中起了多大作用。一个大脑区域的假设功能指导着任务的选择,以检验假设。虽然这些假设和任务通常是基于某个领域的先验知识而选择的,但任务在多大程度上负责创建作为神经回路的假设属性的神经表征呢?
大脑皮层的功能多种多样,几乎涵盖了我们对世界的主观体验的所有方面。这些不同的功能在皮层中是如何组织的?此外,这对于选择身体质量指数应用的皮层位置又意味着什么呢?这些重要的问题将在下一节讨论。
大脑皮层功能定位简史
皮层的定位视图
保罗布罗卡的(1861年)著名的病人谭在额叶损伤后有严重的语言表达障碍,他提供了大脑皮层内功能定位的第一个具体例子(尽管当时他错误地将根本原因归因于纹状体)。概括地说,定位观点认为,人类行为的巨大复杂性可以被分离成特定的功能,每个功能都被定位到大脑皮层的特定区域。布罗卡的结果与约瑟夫一致高尔的(1798)颅相学是一种极端的定位观点,认为大脑皮层的每一部分都有独特的功能。例如,有几块大脑皮层专门负责自尊、仁慈、好斗和模仿。在光谱的另一端,卡尔拉什利(1929)提出了大脑皮层的等电位理论,其中大脑皮层在功能上是无差别的,所有的认知能力都随着大脑皮层损伤的大小而单调下降。这一观点可能受到物种选择的影响(Lashley研究了啮齿类动物,它们的行为主要由皮质下区域介导)。在运动控制中,随着精细运动控制的进化,功能的大脑化增加,使得动物模型的选择至关重要。虽然人类的运动功能可能会被运动皮层的中风所破坏,但啮齿动物的运动执行能力似乎受到的影响要小得多,学习新任务的能力只会下降(Kawai等人,2015年).
网络视图
随着电生理学和精细神经解剖学的出现,简单的定位概念显然是站不住脚的。相反,皮质功能来自许多大脑区域的网络化互动,每个区域都是网络中的一个节点。功能性磁共振成像(fMRI)进一步强调了这一观点,显示了简单任务的多个区域的激活。现在的一个核心问题是,网络中每个节点的区别是什么,以及节点网络如何协同工作来产生复杂的行为?研究人员已经提出了运动系统内的几种组织方案,包括用于控制身体不同部分的节点(安达信和布尼奥2002年),不同类别的行为(Graziano & Aflalo 2007, Kaas & Stepniewska 2016),以及从感知到运动的不同计算阶段。例如,考虑到运动的控制涉及许多元素,例如基于当前需求的目标选择,坐标变换将眼睛坐标中的目标位置转换为身体坐标,规划出实现目标的适当动作序列,并将预期动作转换为肌肉活动的特定序列。这些计算是如何在神经元群体中编码并在大脑皮层中分布的?这个问题可以从动物研究的许多方向来探讨,包括节点的激活和失活。人类BMIs通过记录来自几个皮层区域的神经元群体,结合主观经验的交流,为这个问题提供了一个独特的窗口。
分布式表示
理解一个节点的一个方法是理解它对皮层功能的贡献。在我们实验室的早期实验中,我们研究了超个人空间在NHPs的皮层中是如何表现的。这(法)离开(our prendre conge)在意识到个人以外的空间中起着关键的作用,正如从人类和NHPs的损伤中所证明的那样(福尔摩斯1918, 米什金-昂格莱德1982年, 鲁道夫·巴林特1909年).我们推断,视觉空间将由独立于刺激的视网膜位置的PPC神经元的感受野编码。相反,在NHP记录的单个神经元中,我们发现眼睛位置信号与视网膜刺激位置信号相结合,并且在单个神经元水平上通常没有空间的不变表示(安德森等人,1985年).眼睛位置对视觉感受野的影响近似于单调的眼睛位置信号乘以视觉感受野,我们将这种类型的相互作用称为增益场。当时,三层神经网络和反向传播学习(现代人工智能和机器学习的前身)刚刚开发出来(Rumelhart等人,1986年).我们表明,被训练来表示从视网膜刺激和眼睛位置提取的空间的神经网络形成了相同的分布式表示,其中不变的空间位置只能从神经群体中读出(齐普瑟&安德森1988).眼睛位置增益场并不局限于PPC,在几个皮层区域都有发现(综述见萨利纳斯&蒂尔公司2000年).
随机混合选择性
了解网络中节点贡献的另一个方法是找出不同区域如何编码信息。NHPs的前额叶皮层中的神经元(Rigotti等人,2013年)和大鼠的PPC(Raposo等人,2014年)编码任务变量的随机非线性混合。这随机混合选择性被提出具有在小的神经元群体中表示许多状态的计算优势,这些神经元群体可以容易地用线性解码器(Fusi等人,2016年).此外,有人提出,用高度专门化的神经元处理信息的区域比那些表现出混合选择性的区域具有更低的维度。关联皮层区与初级感觉和运动区的直接比较可以确定当与初级区相比时,关联区是否使用更混合的选择性策略。
人类后顶叶皮层
如上所述,NHPs中相关皮层区域的记录产生了分布在神经元群体中的变量的高维编码的显著发现。随着四肢瘫痪者大脑皮层的记录,主要是PPC的记录,现在变得越来越清楚,在人类中发现了类似的混合变量编码策略。这些表示不是随机混合的,而是具有定义的结构部分混合选择性。在下面的章节中,我们回顾了在人类中,在示例性的关联区域PPC和初级感觉区域中发现的变量的类型和结构初级躯体感觉皮层(S1).
意象
运动想象
想象被认为是大脑皮层回路的内省激活,与这些回路的正常功能重叠。例如,想象的运动会激活参与运动计划和执行的神经元群。在NHPs中,PPC在reach规划和执行期间是活动的,而M1主要在执行阶段活跃(克拉姆蒙德&卡拉斯卡2000, Snyder等人,1997年). 来自人类四肢瘫痪参与者的M1记录显示想象的激活(Hochberg等人,2006年)和企图(科林格等人,2013年, Hochberg等人,2012年)动作。这些激活主要与期望的运动速度有关。
来自人类PPC的记录显示了在计划和执行想象的运动期间的激活(Aflalo等人,2015年) (图2).计划活动与运动的目标相关。目标解码的优点包括:指定目标需要非常少的神经元;解码目标很快(∼200 ms);表征是双向的,一些神经元编码记录半球的对侧肢体,另一些编码同侧肢体。通过直接或象征性的暗示对目标进行编码。控制实验表明,延迟任务中的持续活动确实编码了参与者的意图。NHPs中身体质量指数的早期研究显示了类似的预期目标编码,并被提议为解码和执行运动目标的认知身体质量指数的实例(穆萨拉姆等人,2004年).
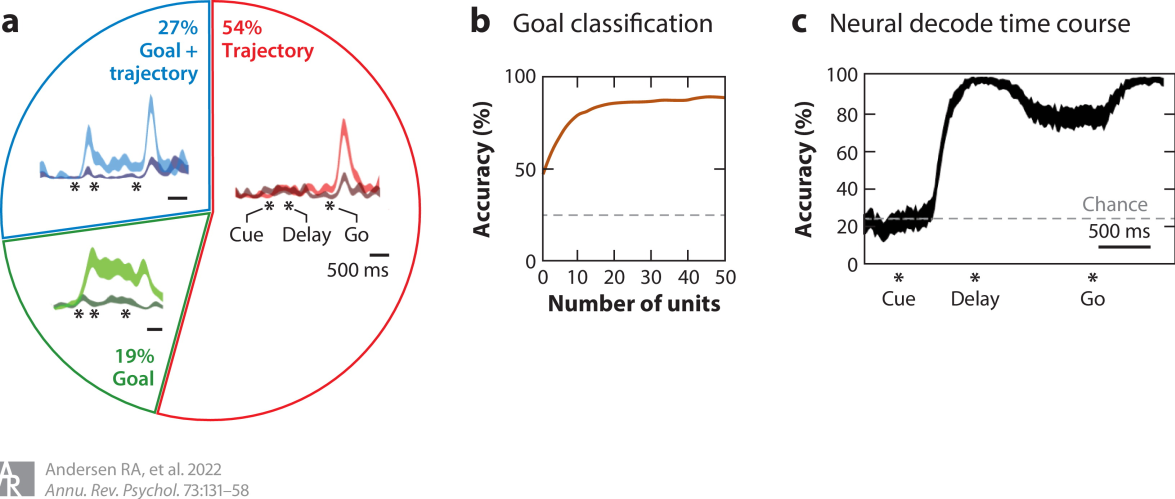
图2人类运动意图的认知编码。(a)PPC神经元用异质混合的神经元编码运动目标和轨迹。饼图显示了不同反应类型在PPC神经元群体中的比例。插图显示了三个代表性示例神经元的活性(平均SEM)。每一个神经元都显示了对目标的响应,引起最大的活动(浅色调)并且响应于相反的目标(暗色调). (b)少量的细胞能够对预期目标进行精确解码。(c)从离线数据计算的神经解码的时间过程证明了在目标呈现的190 ms内对预期目标的准确编码(300 ms滑动窗口,95%置信区间)。图改编自Aflalo等人(2015年)。缩写:PPC,后顶叶皮质;测量的标准误差。
想象的把握
最初的人植入PPC的目标部位是左侧PPC,通过使用功能性磁共振成像(fMRI ),这些部位对于抓握和伸展是活跃的。抓握区位于顶内沟和中央后沟的交界处,可能是大脑皮层的同源物。前顶叶内区在NHPs(康诺利等人,2003年).到达选择性区域在顶上小叶中,并且可能对应于NHPs中的Brodmann区域5d(Cavina-Pratesi等人,2010年).因为很难在NHPs和人类的相关皮质中找到相似性,我们也用一个更中性的解剖学描述符来描述推测的AIP部位,即中央后-顶叶内区域。在一名受试者中选择了位于边缘上回的第二个抓握选择部位。
NHPs中的AIP是抓握回路的一部分,AIP编码可抓握物体的视觉结构,腹侧前运动皮层编码中间感觉和运动成分,M1编码运动成分(Gerbella等人2017年, Schaffelhofer & Scherberger 2016).特定的抓取形状可以从NHPs中的M1解码(巴尔加斯-欧文等人,2010年)和人类(Rastogi等人,2021年, Wodlinger等人,2014年).使用BMIs的M1解码支持机器人手的简单抓取(Hochberg等人,2006年, Wodlinger等人,2014年).从认知身体质量指数的角度来看,AIP中的单个神经元对可以指定整个抓握的物体形状具有选择性(Murata等人,2000年).
在人类AIP中,独立于要抓握的物体的手的想象形状可以被解码(Klaes等人,2015年).视觉线索以及对适当手形的想象来抓住物体激活的AIP。相比之下,听觉线索并没有激活AIP,但是由听觉线索指示的手形产生了同样的想象诱发的激活,使得能够对想象的抓握进行解码。尽管5d区确实对视觉提示有反应,但它的神经元对想象的手形没有选择性。这一结果与NHP记录和人类成像研究的PPC内抓握和伸展加工的分离是一致的。
部分混合选择性
双侧想象运动的发现(Aflalo等人,2015年)和把握活动(Klaes等人,2015年)在人类的AIP中提出了这样一个问题:在这个皮层区域还代表着什么其他的认知变量。在NHP的研究中,使用了在顶叶内沟(IPS)周围的记录和失活,一致的偏好效应物在嘴唇中有更多的眼球运动,在顶骨延伸区域有更多的延伸运动,在AIP中有更多的抓握运动(安达信和布尼奥2002年).早期对人类的功能磁共振成像研究呈现出更多的混合结果,一些研究报告了效应物的分离(阿斯塔菲耶夫等人,2003年, 康诺利等人,2003年, Culham等人,2003年, Gallivan等人,2011年, 普拉多等人,2005年)和其他报道了效应器的混合(Beurze等人,2009年, Heed等人,2011年, 欣克利等人,2007年).用于触及和抓取的效应器表示是双手的(Gallivan等人,2013年),与人类AIP的记录一致(Aflalo等人,2015年).
一项研究旨在检查三个变量的表征程度:尝试或想象的运动,同侧或对侧身体侧,以及肩和手的效应器(张等2017) (图3).这些变量的八种可能组合被用作指示参与者运动的线索。这些提示以文本指令的形式呈现,之后是延迟提示和继续提示以产生反应。例如,一种试验类型是文本“想象右手”,它指示参与者想象(在他们的脑海中想象运动)挤压他们的右手。令人惊讶的是,所有这些变量都在AIP人群中发现,并且是混合的,单个神经元经常对一个以上的变量做出反应。然而,这种混合不是随机的,而是有结构的。如果身体的一侧被描绘,另一侧也很可能被描绘;尝试或想象运动的认知策略也是如此。然而,对于效应器,单个神经元的混合是随机的。例如,代表手部运动的神经元很可能不代表肩部运动。虽然只有不同的效应器表现出强烈的分离,但一个任务中三个变量的所有组合都可以被解码。
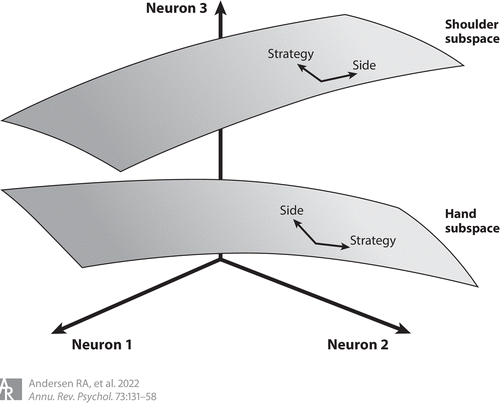
图3部分混合选择性的示意图。PPC以结构化的方式对多个运动变量进行编码。身体运动(手或肩)被编码在不同的子空间(灰色飞机).其他运动变量(身体一侧,左对右;和运动策略,想象与尝试的运动)被编码在共享的子空间中,在它们各自的身体运动中组织。为了说明的目的,示出了三个神经元的运动平面,但是这些运动平面嵌入在记录的群体的整个空间内。图改编自舍尔伯格(2017年)。缩写:PPC,后顶叶皮质。
由于统计结构,这种混合被称为部分混合选择性。这种结构中的相关性的可能优点是能够在身体每一侧的相同效应器之间转移学习(Amemiya等人,2010年)并使用运动想象来改善运动执行(迪克斯坦&德意志2007).效应器中的隔离可能会减少移动身体不同部分所涉及的计算干扰。
一个重要的问题是,部分混合结构是否跨上下文维护。一个强有力的和特别重要的测试身体质量指数视角是开环训练的结构在闭环大脑控制期间是否保持不变。发现不同运动状态的表示在很大程度上保持在两种状态之间(张等2020).